А.Д. Строганов (Московский государственный университет им. М.В. Ломоносова)
Среди позвоночных животных рыбы являются бесспорными чемпионами по разнообразию форм, окраски, адаптационных механизмов для приспособления к многообразию условий окружающей среды. Многие виды рыб являются объектами товарного рыбоводства, а еще большее количество – излюбленные объекты аквариумистики. Декоративное рыбоводство широко использует достижения ихтиологии и промышленного рыборазведения в области физиологии, кормления, лечения рыб. Так, например, вторая половина прошлого века характеризовалась увеличением количества разводимых в аквариумах видов рыб во многом за счет применения методов гормональной стимуляции созревания половых продуктов, разработке которых в нашей стране было уделено огромное внимание исследователей при создании технологий промышленного разведения осетровых и растительноядных рыб (Гербильский, Исаев, Кожин и др., 1963; Макеева, Веригин, 1971).
Но несправедливым было бы не отметить, что велика и доля аквариумистики в процессе познания особенностей развития, роста, поведения рыб. Разработанные и усовершенствованные методы содержания и разведения рыб позволили получить бесценный материал, например, для изучения эмбрионально-личиночного развития различных экологических групп рыб и, особенно, различного уровня дыхательных приспособлений у эмбрионов и личинок.
Вот, например, три вида рыб - нотобранхиус, гуппи и макропод. Из всего многообразия аквариумных рыб они представляют различные экологические группы (Крыжановский, 1948) и демонстрируют поразительное разнообразие дыхательных адаптаций в раннем развитии (Соин, 1968).
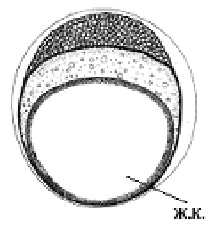
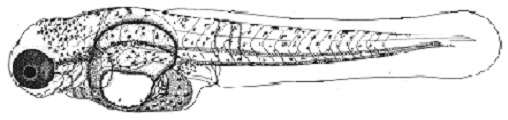
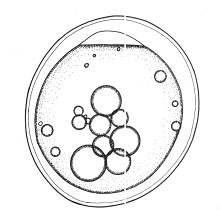
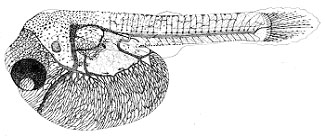
Развитие макропода проходит в идеальных, с точки зрения насыщения кислородом, условиях. Эмбрионы и личинки за счет огромной жировой капли в желточном мешке (рис.1, а) обладают положительной плавучестью в стоячей воде (в отличие от рыб морских, очень редко встречающаяся у пресноводных рыб адаптация). Как и у большинства пелагофилов, эмбриональные органы дыхания отсутствуют, набор дыхательных механизмов минимальный (до вылупления только нервно-мышечная моторика), вылупление из икры на ранних стадиях слабосформированной предличинки (без кровеносной системы, систем пищеварения и дыхания) (рис.1, в).
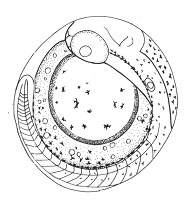
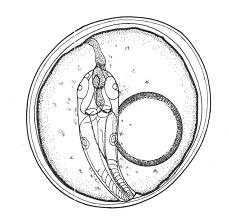
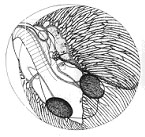
Развитие нотобранхиуса проходит в разнообразных по содержанию кислорода условиях, но попадание в условия осушения привело к необходимости формирования эмбрионом третьей яйцевой оболочки – экстраэмбриональной мембраны, с одной стороны, предохраняющей икринку от высыхания, но, с другой стороны, являющейся дополнительным препятствием для проникновения к телу эмбриона кислорода. Это явилось одной из основных причин того, что нотобранхиус во время развития под оболочкой обладает целым спектром сменяющих друг друга дыхательных приспособлений и механизмов (движения тела, подвижность грудных плавников и жаберно-челюстного аппарата; клетки крови и гемоглобин появляются практически одновременно с формированием сердца и сосудов; развивается эмбриональный орган дыхания - сосудистая сеть на желточном мешке). И даже более того, темп развития эмбриона нотобранхиуса, по сравнению с другими рыбами, значительно снижен, что также позволяет уменьшить кислородные потребности.
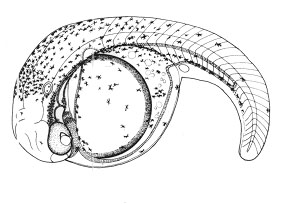
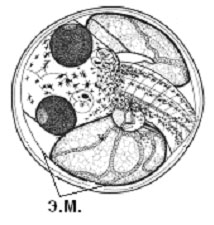
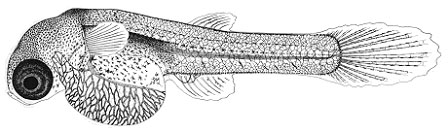
Из всех трех видов эмбрионы гуппи, развиваясь внутри полости тела самки, находятся в самых суровых с точки зрения потребления кислорода условиях. И, как следствие, обладают наиболее сильно развитыми эмбриональными органами дыхания – мощная сеть кровеносных сосудов покрывает не только желточный мешок, но развивается и на теле. Более того, желточный мешок образует покрытые сетью сосудов выросты, охватывающие голову эмбриона (т.н. «ошейник») и увеличивающие площадь дыхательной поверхности.
Таким образом, степень развития эмбриональных органов дыхания и дыхательных механизмов находится в зависимости от кислородных условий среды и, в значительной степени, различается у рыб, представляющих различные экологические группы.
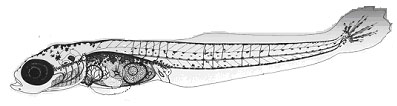
Необходимо также отметить и другую корреляцию: если рассматривать раннее развитие до перехода на смешанное питание, то, с энергетической точки зрения, можно сказать, что мы рассматриваем практически замкнутую систему эмбрион – желток. И можно ожидать, следуя закону сохранения энергии, дополнительные безвозвратные потери питательных веществ желточного мешка на формирование эмбриональных органов дыхания и механизмов. Данное положение вполне подтверждается сравнительными оценками эффективности использования метаболизированной энергии желточного мешка у представителей разных экологических групп. Оказалось, что у пелагофила с минимумом дыхательных приспособлений – трески этот показатель выше, чем у представляющего литофильную группу атлантического лосося (Строганов, Новиков, 2001).
Таким образом, из вышеизложенного можно сделать заключение, что изучение особенностей эмбрионально-личиночного развития рыб способствует не только развитию фундаментального знания, но также имеет большое практическое значение, так как позволяет осознанно подбирать условия развития (кислородные, температурные и др.), успешно преодолевать «критические стадии развития», успешно проводить вылупление и, что может быть еще более важно, повышать выживаемость при переходе на экзогенное питание, что, в конечном счете, улучшает экономические показатели работы рыбоводных хозяйств.
|
|
|
|
|
| ||
| Рис. 1. Эмбрионально-личиночное развитие макропода (Macropodus opercularis) Ж.К. – жировая капля а – поздняя морула б – начало обособления хвостового отдела от желточного мешка в – эмбрион на стадии вылупления г – переход на смешанное питание | ||
|
|
|
|
|
| ||
|
Рис. 2. Эмбрионально-личиночное развитие нотобранхиуса (Nothobranchius guntheri) | ||
|
|
|
|
|
| ||
|
Рис. 3. Эмбрионально-личиночное развитие гуппи (Poecilia reticulata) | ||
|
|
| Рис. 4. Эмбриональная моторика литофилов (а – атлантический лосось) и пелагофилов (б – треска) от оплодотворения до вылупления 1 – цитоплазматическая моторика; 2 – подвижность тела зародыша; 3 – пульсация сердца; 4 – подвижность грудных плавников; 5 - подвижность жаберно-челюстного аппарата. |
Литература:
- Авни А.А. 1974. Эколого-морфологические особенности развития некоторых тропических карпозубых (Cyprinodontoidei) и лабиринтовых (Anabantoidei) рыб. Дисс. канд. биол. наук. М.: МГУ. 144 с.
- Гербильский Н.Л., Исаев А.И. Научные основы, направления развития и районирование осетрового хозяйства в водах СССР. В кн.: Осетровое хозяйство в водоемах СССР. Изд-во АН СССР. М.: 1963, с. 5-19.
- Кожин Н.И., Гербильский Н.Л., Казанский Б.Н. Биотехника разведения осетровых и принципиальная схема осетрового рыбоводного завода. В кн.: Осетровое хозяйство в водоемах СССР. М.: АН СССР, 1963, с.29-34.
- Крыжановский С.Г. 1948. Экологические группы и закономерности их развития// Изв. ТИНРО. Т.27. С.3-114.
- Макеева А.П., Веригин Б.В. 1971. Метод гипофизарных инъекций в практике разведения толстолобиков и белого амура. Вопр. Ихтиологии. Т.11. Вып.2. С.217-231.
- Соин С.Г. 1968. Приспособительные особенности развития рыб. М.: МГУ. 89с.
- Строганов А.Н., Новиков Г.Г. Особенности становления дыхательной функции в раннем онтогенезе костистых рыб. В кн.: «Экологические проблемы онтогенеза рыб: физиолого-биохимические аспекты», М.: МГУ. 2001. С.20-30.
 Морские рыбки
Морские рыбки
 Пресноводные рыбки
Пресноводные рыбки
 Террариумные животные
Террариумные животные
 Аквариумы и тумбы
Аквариумы и тумбы Каталог товаров
Каталог товаров Терарриумы
Терарриумы