А.Н. Строганов(1), Г.Г. Новиков(2)
(Московский государственный университет им. М.В. Ломоносова (1), Беломорская биологическая станция (2))
Изучение закономерностей роста и метаболизма развивающихся зародышей рыб имеет важное значение как для понимания становления механизмов адаптации к меняющимся условиям среды, включая формирование внутривидовой разнокачественности особей и популяций, так и для совершенствования методов искусственного воспроизводства рыб путем создания биотехнических режимов, обеспечивающих максимально эффективное использование потенциальных возможностей вида. Несмотря на то, что первые исследования в этом направлении были выполнены еще в конце позапрошлого - начале прошлого столетия, многие проблемы остаются недостаточно изученными. И одна из них – это степень сопряженности процессов в условиях температурного воздействия. Согласно сложившимся к середине прошлого века представлениям наблюдается «зеркальный» характер (Новиков, 2000) взаимосвязи процессов роста организма и утилизации желтка, высокая степень сопряженности процессов роста и морфологического развития (рис. 1).
Однако проведенные на кафедре ихтиологии МГУ экспериментальные работы на эмбрионах и личинках рыб, представляющих различные систематические и экологические группы (треска, пинагор, радужная форель, семга, белорыбица и др.), показали, что особенности действия температуры на процессы трансформации вещества и энергии в раннем онтогенезе рыб имеют более сложный характер, результатом чего являются, например, более крупные размеры эмбрионов на стадии вылупления (рис. 2) и личинок на стадии перехода на смешанное питание (рис. 3) при развитии при более низких значениях температуры (естественно, не выходящих за пределы зоны оптимальных значений).
Зная закономерности влияния факторов среды, в частности температуры, на характер роста, эффективность использования запасных веществ желтка и динамику энергетических затрат в процессе развития рыб, на ранних этапах онтогенеза появляется возможность определения оптимальных значений факторов среды и регуляции развития, т.е. управления развитием и ростом. Это необходимо с целью получения особей не только в заранее планируемые сроки, но и с заданными морфо-функциональными параметрами.
Так, использование модифицированного для эмбрионально-личиночного развития балансового уравнения Г.Г. Винберга (Винберг, 1956) и коэффициентов использования запасных веществ В.С. Ивлева (Ивлев, 1939, 1954) дало возможность провести сопоставление показателей эффективности расходования запасных веществ, например, у эмбрионов пинагора (табл. 1) (Новиков, Куфтина,1988; Строганов, 1987). Полученные значения позволяют достаточно четко определить наиболее оптимальные значения температур для этого вида.
Таблица 1. Эффективность использования метаболизированной энергии резорбированных запасных веществ на рост ("К2") у пинагора (за время развития от оплодотворения до вылупления).
|
Условия развития |
"К2" |
| 6,5 оС 24%о | 0,41 |
| 8,5 оС 24%о | 0,56 |
| 10,0 оС 24%о | 0,42 |
| 12,1 оС 24%о | 0,26 |
Таким образом, мы наблюдаем специфическое действие температур на процессы роста, развития, резорбции желтка, дыхания и др. С одной стороны, это приводит к разнокачественности морфо-физиологических параметров в группах рыб, а, с другой стороны, дает инструмент для управления и регуляции процессами трансформации вещества и энергии в раннем онтогенезе рыб. С практической точки зрения появляется возможность разработки режимов управления абиотическими факторами, например температурой, распределения энергетических затрат в раннем развитии рыб, что позволяет получать в планируемое время молодь рыб с заданными морфо-физиологическими характеристиками. Интересно отметить, что эти методики, все более широко используемые в товарном рыбоводстве, могут найти свое применение и в аквариумном рыбоводстве, так как экономический эффект от повышения выживаемости и ускорения темпов развития и роста для аквариумных рыб будет значительно выше в связи с их высокой стоимостью и относительно низкой плодовитостью.
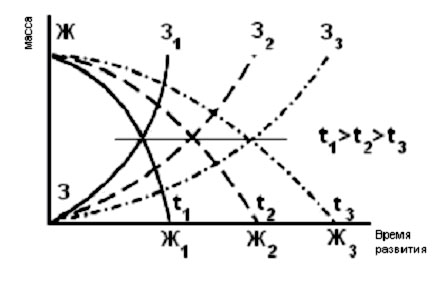
Рис.1. Схема, иллюстрирующая представления о «зеркальном» характере взаимосвязи процессов роста зародыша рыб и резорбции желтка при разных температурах развития
З - масса зародыша; Ж - масса желтка; tоС - температура развития
(по Новикову, 2000).
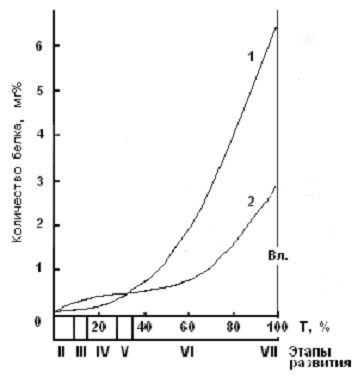
Рис. 2. Рост массы тела зародыша пинагора (от оплодотворения до вылупления) при разных температурах развития (1 – 8,0оС; 2 – 11,6оС).
Вл. – вылупление из икры (по Новикову, 2000).
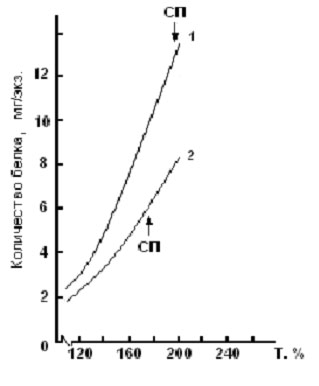
Рис. 3. Рост массы тела предличинок атлантического лосося от вылупления до перехода на смешанное питание (СП) при разных температурах развития (1 – 2,9оС; 2 – 8,3оС) (по Новикову, 2000).
Литература:
- Винберг Г.Г. 1956. Интенсивность обмена и пищевые потребности рыб. Минск: БГУ. 254 с.
- Ивлев В.С. 1939. Энергетический баланс карпов// Зоол.журн. Т.18, вып.2. С.303-318.
- Ивлев В.С. 1954. Зависимость интенсивности обмена у рыб от веса их тела// Физиол. журн. Т.40, № 6. С.717-721.
- Новиков Г.Г. Рост и энергетика развития костистых рыб в раннем онтогенезе. М.: Эдиториал УРСС. 2000. 296 с.
- Новиков Г.Г., Куфтина Н.Д. 1988. Особенности роста зародыша и закономерности утилизации запасного белка в желтке в раннем онтогенезе пинагора при разных температурах развития. – 1.Динамика накопления белка в теле зародыша и резорбция белковой части желтка// Вестник МГУ. С.7-12.
- Строганов А.Н. 1987. Закономерности изменения потребления кислорода и особенности энергетического обмена у некоторых видов рыб на ранних этапах онтогенеза при различных значениях абиотических факторов. Дисс. канд. биол. наук. М.: МГУ. 151 с.
 Морские рыбки
Морские рыбки
 Пресноводные рыбки
Пресноводные рыбки
 Террариумные животные
Террариумные животные
 Аквариумы и тумбы
Аквариумы и тумбы Каталог товаров
Каталог товаров Терарриумы
Терарриумы