А.Н. Строганов
Московский государственный университет имени М. В. Ломоносова
Кета - Oncorhynchus keta (Walbaum, 1792) - представитель рода тихоокеанских (или дальневосточных) лососей, обитает в северной части Тихого океана, на нерест поднимается в многочисленные реки азиатского и американского побережий. На азиатском континенте кета для нереста использует реки от Лены, Колымы и Индигирки на севере, до реки Тумнминцзян (Туманган, Тумень-Ула, Туманная) на юге Приморья (Кузнецов, 1937; Берг, 1948; Андрияшев, 1954). Нерестится кета и в реках Курильских островов, на острове Хоккайдо и в северной части о. Хонсю. На американском континенте кета встречается от Берингова пролива до залива Монтеррей (штат Калифорния) (Gilbert, 1924; Ricker, 1954).
Для размножения кета использует как крупнейшие речные системы, такие как Амур, Камчатка, Анадырь, Юкон, Макензи, так и небольшие речки. Если сравнивать уловы кеты в северной Америке и Азии, то преимущество будет сильно сдвинуто в сторону азиатского побережья.
У кеты описано несколько внутривидовых форм. Наиболее известными является летняя и осенняя форма кеты (Берг, 1934, 1948, 1955; Кузнецов, 1937). Необходимо отметить, что описанные формы отличаются от одноименных у атлантического лосося – сёмги, так как осенняя кета, в отличие от сёмги, нерестится в том же году. Осенняя кета входит в реки позже летней и, соответственно, нерестится также позже. Различна и морфология нерестилищ летней и осенней кеты. Летняя кета для размножения выбирает участки с галечниковым дном, расположенные, как правило, в конце плесового участка реки, перед перекатом, где скорость подруслового потока воды достигает максимальных значений. Температура воды во время нереста летней кеты составляет около 12-130С, температура грунта, куда закапывается икра, может быть несколько выше. По мере наступления осени и зимы температура воды и, соответственно, температура грунта постепенно снижаются и в январе, феврале температура в гнездах летней кеты может составлять до 0,10С. В холодные зимы нерестилища летней кеты часто промерзают, что сопровождается гибелью икры и предличинок.
Нерестилища осенней кеты располагаются в местах выхода грунтовых вод. Самка осенней кеты сооружает гнездо в месте выхода ключа, или, вблизи действующего источника. Большая часть нерестилищ осенней кеты располагается именно в ключевых протоках и ключевых затонах. Во время нереста осенней кеты в сентябре температура воды составляет около 7-80С. По сравнению с икрой летней кеты, икра осенней кеты развивается в стабильных температурных и кислородных условиях, в зимние месяцы температура в гнездах никогда не опускается ниже 2-30С. Поэтому даже в морозы до –500С (Магаданская обл.) нерестилища кеты не покрываются льдом (Волобуев, 1983; Волобуев и др., 1990). В связи с более стабильными температурными и кислородными условиями суровые зимы в меньшей степени влияют на воспроизводство осенней кеты.
Приуроченность обеих форм кеты к разным частям ареала объясняется, скорее всего, геологическими и ледниковыми процессами, происходившими в четвертичный период. Во время трансгрессий (отступлений) моря во времена оледенения значительные пространства морского дна поднимались, становясь сушей. Во времена наибольшей трансгрессии моря Приморье, Сахалин, Курилы и Амур являлись одной крупнейшей водной системой – Палеоамуром, что повлияло на особенности расселения кеты в регионе.
Таким образом, на большей части ареала кета предпочитает для нереста участки рек, где имеет место устойчивый дебет грунтовых вод на обширных участках (Евзеров, 1986; Путивкин, Морозов, 1988, Волобуев и др., 1990). Сокращение площадей – участков с выходами грунтовых вод, в результате человеческой деятельности приводит к довольно сильному сокращению стад осенне-нерестующей кеты. Сокращение участков вызвано, как правило, вырубкой лесов на берегах рек, строительством на берегах, и, естественно, гидростроительством, приводящим к дисбалансу уровня подземных источников и воды в реке. В то же время, нерестилища летней кеты также подвержены влиянию человека, причем даже в большей степени, чем нерестилища кеты осенней. Это связано как с указанными выше факторами, так и очень сильным заилением грунта нерестилищ летней кеты, которое может быть следствием и вырубок, и строительства, и собственно загрязнения, а также нерациональным ведением сельскохозяйственного производства на берегах рек или вблизи них.
Средняя длина половозрелых особей кеты составляет 60-70 см, вес 3-4,5 кг; рекордные экземпляры вырастают более метра и весят свыше 14 кг. Плодовитость в среднем 3200-4200 икринок. Диаметр набухшей икры в среднем 7-8 мм, масса от 180 до 300 мг. Для развития кеты считается благоприятным содержание кислорода от 4,5 до 7-8 мг/л. Скорость проточности (фильтрации) воды в гнездах разных нерестилищ колеблется в среднем от 11 до 36,5 м/час.
Массовое вылупление из икринок предличинок кеты в разных популяциях происходит при разном количестве градусо-дней. Среднее значение соответствует 520-530 градусо-дням при длине предличинки 22-23 мм. При длине 24-26 мм предличинки переходят к подвижному состоянию. Переход к смешанному питанию начинается при достижении длины 30-32 мм и массы 230-255 мг и остатке желтка 27-35% (возраст от 612 до 760 градусо-дней). Скат молоди из небольших рек начинается на этапе смешанного питания при средней длине 38 мм и массе 335 мг. Из более крупных рек молодь мигрирует при длине 35-45 мм и массе 333-600 мг. В ряде случае молодь может задерживаться в реке до 3-4 мес., достигая длины 5-7 см. В соответствии с нормативной рыбоводной документацией выпуск молоди с рыбзаводов осуществляется при массе от 0,3 до 2,0 г.
Целью настоящих работ является исследование особенностей обменных процессов и роста у кеты на ранних стадиях онтогенеза при воздействии, главным образом, температурного фактора. Поводом для проведение работ является развитие в мире биотехнологий культивирования рыб и беспозвоночных с применением интенсивных методик, способствующих повышению экономических показателей и снижению себестоимости продукции. Наиболее ярким примером является увеличение производства товарной семги в мире до 1,24 млн. т (стоимость 4,77 млн. USD) (FAO, 2005), что явилось результатом применения в раннем развитии высоких температурных режимов, с которыми в природе данный объект (на сходных стадиях развития) не встречается, но, как было выяснено в ходе научно-исследовательских работ, прекрасно переносит, развивается и растет, что дало возможность в 3-4 раза сократить период наступления смолтификации (серебрения) и создать экономически выгодные технологии культивирования семги. При этом, убыточность культивирования семги при природных, естественных температурных режимах спровоцировала идущий в настоящее время процесс по сворачиванию деятельности и закрытию лососевых рыбоводных заводов в Мурманской области[1].
В качестве объекта исследований использовались предличинки кеты перед началом перехода на эндогенное питание, доставленные с ЛРЗ «Лазовой» (Макаровский р-н, Сахалинская обл.). Наиболее полно рассматривалось действие температурного фактора и солености на рост, питание и обмен кеты в раннем онтогенезе. Предличинки были разделены на группы и рассажены в отдельные емкости, размещенные в помещениях холодной комнаты и бытовых холодильников. Объемы емкостей, интенсивность аэрации и фильтрации воды (а также, соответствующее оборудование) выбирались исходя из возраста, размеров и численности исследуемых объектов. Сведения о режимах воздействия температуры и солености, а также времени проведения экспериментальных работ в отдельных группах представлены в табл. 1. Испытание различными концентрациями солености производилось после начала процесса смолтификации у молоди кеты.
Для контроля действия факторов на предличинок и мальков кеты контролировали выживаемость, проводили измерения длины (мм) и веса (г).
По наступлении этапа смешанного питания предличинкам в различных группах давали стандартный стартовый лососевый корм, использовавшийся в 2006 г. на ЛРЗ «Лазовой». Режим питания и объем различались в различных температурных вариантах. В связи с отсутствием кормов более крупных фракций по мере роста малькам давали мелкого, а затем и крупного мотыля.
Таблица 1. Схема опытов.
|
Номер группы |
Темп-ра, 0С |
Соленость, %о |
Период |
|
Гр.1 |
6-8 |
0 |
17.04.06 - 29.09.06 |
|
Гр.1* |
7-8 |
постепенное увелич. от 6 до 10 |
24.07.06 - 29.09.06 |
|
Гр.2 |
6-15 |
постепенное увелич. от 0 до 30 |
25.04.06 - 10.10.06 |
|
Гр.2* |
7-8 |
постепенное увелич. от 6 до 10 |
20.07.06 - 29.09.06 |
|
Гр.3 |
10-12 |
0 |
18.04.06 - 10.12.06 |
|
Гр.3* |
10-12 |
постепенное увелич. от 4 до 8 |
27.07.06 - 13.09.06 |
|
Гр.4 |
13-15 (17) |
постепенное увелич. от 0 до 30 |
30.04.06 - 05.10.06 |
|
Гр. 7 |
15-16 |
0 |
30.04.06 - 08.06.06 (объед. с гр.4.) |
|
Гр. 8 |
постепенное увелич. от 13 до 19 |
0 |
05.05.06 -07.05.06 |
У предличинок и мальков кеты на отдельных стадиях развития периодически проводили измерение 12 морфометрических признаков (рис. 1) таких, как полная длина тела (L), длина тела до выемки в хвостовом плавнике (длина по Смитту) (Ls), длина тела без учета длины хвостового плавника (l), длина головы (C), высота головы (Ch), наибольшая высота тела (H), высота хвостового стебля (h), антедорсальное расстояние (AD), постдорсальное расстояние (PD), длина хвостового отдела (Cl), диаметр глаза (o) и интерорбитальное расстояние (io).
Все промеры производились с точностью до 1 мм (отдельные до 0,5 мм).
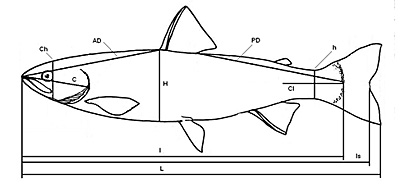
Рис. 1. Схема промеров кеты.
Обозначения: L - полная длина тела; ls - длина по Смиту; l - длина тела без учета длины хвостового плавника; C - длина головы; Ch - высота головы; H - наибольшая высота тела; h - высота хвостового стебля; AD - антедорсальное расстояние; PD - постдорсальное расстояние; Cl - длина хвостового отдела.
Исследовали особенности влияния температуры (в диапазоне от 5 до 18-19оС) и солености на выживаемость и рост (масса) в раннем онтогенезе кеты. Обнаружено, что температуры в диапазоне от 5 до 16 оС не оказывали влияние на выживаемость личинок и мальков кеты (рис. 2), не обнаружено и явного действия температуры на пластические признаки кеты (табл. 2). Повышение и понижение температур с достаточно высокой скоростью (до 5 -7оС в сутки не оказывало влияния на выживаемость. При этом медленное 1-2оС в сутки повышение температуры сверх 16оС вызвало 100% гибель личинок кеты в течение 1 сут. Верхний температурный порог устойчивости личинок кеты в наших опытах составлял 18оС. Учитывая то, что при температурах до 16оС не отмечено снижения выживаемости, а также характерных для крайних значений зон толерантности ухудшения питания, снижения приростов, следует температурный диапазон 16,5-17,5оС определить, как зону верхних значений толерантности к температурному фактору.
Таким образом, в ходе проведенных работ выявлено, что у личинок кеты (практически перед переходом в мальковое состояние) очень узкая зона верхних предпороговых значений температуры. Такая ситуация для аквакультуры представляет определенную опасность, так как относительно небольшое колебание температуры в зоне предпороговых значений может инициировать возникновение необратимых тератогенных процессов и гибель (в опыте снижение температуры от 18 до 14оС не препятствовало летальному исходу). Что касается рекомендаций для производственных предприятий использований температур на границе зоны устойчивости, то необходимо учесть, что полученные нами значения пороговых и предпороговых значений температуры получены в экспериментальных условиях, когда исключался аддитивный эффект других факторов. Содержание кислорода при всех исследованных температурах находилось всегда в пределах 100%, выбранные значения температуры практически не изменялись ни во времени ни в пространстве, чего невозможно достичь в производственных условиях ни в инкубаторах, ни в бассейнах, ни в питомниках, ни, тем более, в прудах, где всегда наблюдаются флюктуации температурного и кислородного факторов и в течение суток, и в толще воды. В связи с вышеизложенным на практике, принимая чрезвычайно узкий диапазон предпороговых значений и необратимость проходящих в организме изменений, ведущих к летальному исходу, следует принимать верхнюю температуру для подращивания личинок и выращивания молоди ниже на 2оС, то есть в пределах 14оС.
Возвращаясь к результатам проведенных нами работ, следует отметить, что наблюдалось выраженное позитивное влияние высоких температур (в зоне толерантности) на питание и рост личинок и мальков кеты. Повышение температуры развития от 5 до 15-16 оС в пресной воде инициировало повышение темпа роста (рис. 3-7). Важным фактором для производства является поедаемость корма. Проведенные наблюдения поисковой активности показали, что наиболее благоприятным в проведенных опытах являлся диапазон в пределах 12-16 оС. Пищевая и поисковая активность в производственных условиях имеют особенное значение, так как кормление производится искусственными неживыми кормами, непоедание которых, даже в течение относительно небольшого периода, приводит к загрязнению воды и непроизводительному увеличению расхода кормов, что значительно ухудшает экономические показатели при высокой производственной мощности предприятия. В ходе выполненных экспериментальных работ увеличение температур развития выше 11оС на разных периодах развития вне зависимости от того содержалась кета ранее при высоких температурах или быстро переведена из низких температурных режимов (3-6оС) поисковая и пищевая активность при кормлении искусственным стартовым лососевым кормом (идентичным использовавшемуся в сезон 2006 г. на ЛРЗ «Лазовой») резко возрастала, что приводило к практически 100% поедаемости задаваемого корма при многоразовом режиме кормления. Причем, наблюдалась определенная дифференциация молоди по способу питания: либо с поверхности воды, либо со дна. Таким образом, высокие температуры способствовали более равномерному и полному поеданию кормов молодью.
Все вышеизложенное касается пресноводного периода жизни молоди кеты. Нами не производились работы по исследованию осморегуляторной ситемы кеты в раннем развитии – в период до и в ходе смолтификации. Однако, в связи с тем, что молодь была подразделена на большое количество групп с различными температурными и соленостными условиями, где наибольшее внимание было уделено исследованию динамики роста, то сравнение особенностей изменения массы (известно, что при смолтификации нахождение в пресной воде вызывает снижение темпа роста (рис. 3-6), голодание и похудение) показало, что вне зависимости от температуры развития во всех вариантах опыта, как при температурах 7-8, 10-11 и 13-14оС, практически в тот же календарный период (конец июля) отмечено на фоне падения пищевой активности снижение темпа роста и падение средней массы у мальков кеты. Подтверждением того, что именно содержание в пресной воде было повреждающим фактором явилось то, что кормовая активность и темп роста восстановились после перевода мальков в соленую воду и интенсивного увеличения солености.
Таким образом, проведенные работы позволили получить результаты, применение которых (особенно на производственных предприятиях с возможностью управления абиотическими факторами) позволит выбирать наиболее оптимальные режимы воздействия факторов среды с целью получения крупной молоди с высокой выживаемостью в короткие срок и с низкими материальными затратами (рис. 8, 9). Применительно к ЛРЗ «Лазовой» с целью снижения затрат при подращивании личинок и выращивании молоди в прудах возможно длительное выдерживание предличинок при низких температурах и даже отсрочка дачи корма личинкам на этапе смешанного питания на фоне низких температур с целью выжидания периода паводков и повышения естественных температур выше 10оС. После этого молодь может быть переведена в пруды с интенсивным кормлением и высокими температурами (12-13оС) и выращена до нормативных значений массы за 10-15 дней.
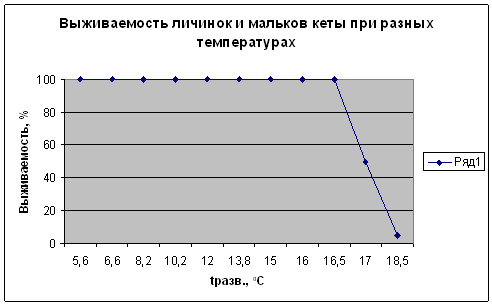
Рис. 2. Выживаемость кеты при разных температурах развития
Таблица 2. Морфометрические характеристики кеты в раннем развитии.
|
№ группы |
Дата |
L, mm |
Ls, mm |
l, mm |
AD, mm |
PD, mm |
Cl, mm |
O, mm |
C, mm |
HC, mm |
H, mm |
h, mm |
io, mm |
|
гр.1 |
10.07.06 |
49 |
46 |
42 |
19 |
17 |
8,5 |
2,6 |
11,5 |
6 |
7,5 |
|
4,5 |
|
гр.2 |
25.04.06 |
36,5 |
34 |
32 |
17 |
12 |
10 |
2 |
7 |
3,5 |
6,5 |
1,5 |
|
|
гр.2 |
25.04.06 |
34 |
32 |
29 |
16 |
11 |
8 |
2 |
6,5 |
3 |
6 |
1,5 |
|
|
гр.2 |
13.07.06 |
59 |
54 |
49 |
24 |
20 |
|
2,5 |
13 |
7,5 |
8 |
3,5 |
5 |
|
гр.2 |
14.07.06 |
65 |
61 |
54 |
25 |
21 |
12 |
4,5 |
14,5 |
9 |
8,5 |
6,5 |
6 |
|
гр.2 |
05.10.06 |
82 |
73 |
70 |
35 |
27 |
13 |
6 |
20 |
14 |
18 |
4 |
8 |
|
гр.3* |
04.09.06 |
62 |
55 |
50 |
25 |
20 |
9 |
4 |
13 |
8 |
9 |
4 |
6 |
|
гр.3* |
04.09.06 |
67 |
63 |
56 |
30 |
25 |
13 |
4 |
15 |
11 |
12 |
5 |
8 |
|
гр.4 |
13.07.06 |
70 |
62 |
59 |
29 |
25,5 |
12 |
4 |
16 |
9,5 |
10,5 |
5,0 |
7,5 |
|
гр.4 |
05.10.06 |
99 |
94 |
85 |
38 |
36 |
18 |
6 |
23 |
13 |
17 |
6 |
8 |
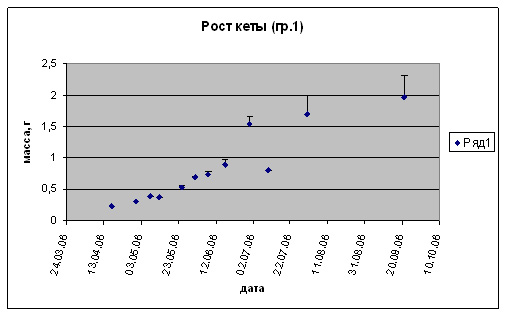
Рис. 3. Рост предличинок, личинок и мальков кеты.
Группа 1 (t разв 6-8 0С S= 0%о.)
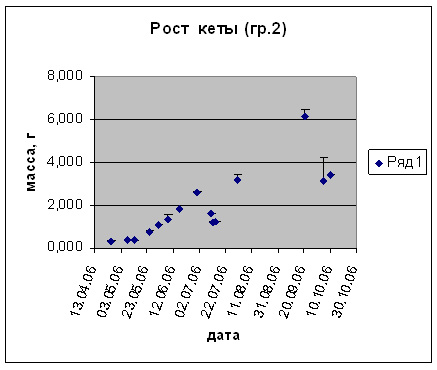
Рис. 4. Рост предличинок, личинок и мальков кеты.
Группа 2 (t разв 6-15 0С, S увелич. от 0 до 30%.)
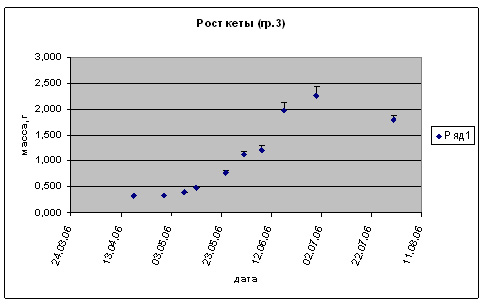
Рис. 5. Рост предличинок, личинок и мальков кеты.
Группа 3 (t разв 10-12 0С, S= 0%.)
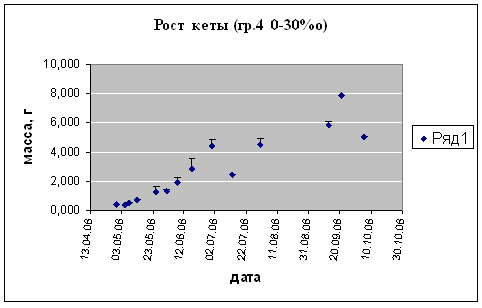
Рис. 6. Рост предличинок, личинок и мальков кеты.
Группа 4 (t разв13-15 0С (170С). S= увелич. от 0 до 30%о.)
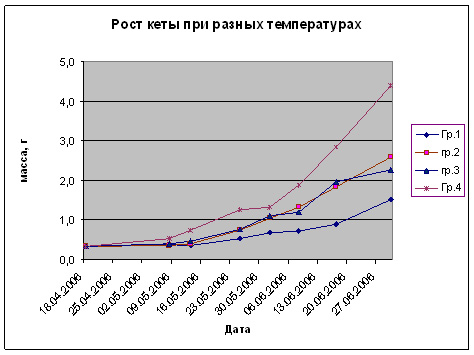
Рис. 7. Влияние температуры развития на рост предличинок, личинок и мальков кеты.

Рис. 8. Рост кеты при разных температурах (возраст 20 сут. от начала питания)
А - t разв. 13-150С, Б - t разв. 6-8 0С
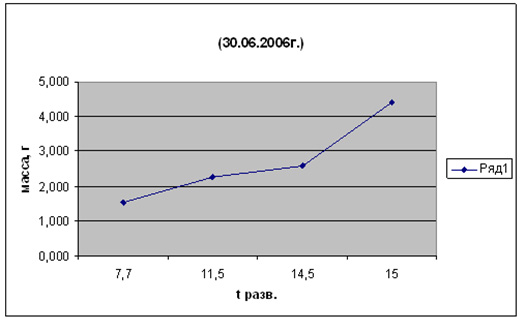
Рис. 9. Влияние температуры развития на рост кеты
Одним из направлений аквакультуры, предполагающих возможность использования модифицированных (акселерирующих) режимов воздействия факторов, являются модульные технологии, где на фоне резкого снижения стоимости работ по основному строительству, появляются широкие возможности снижения себестоимости производимой продукции путем стимуляции процессов роста и развития культивируемого объекта.
Рыбоводный завод модульного типа представляет собой комплекс унифицированных конструкций с профилированием по отдельным этапам рыбоводного процесса: инкубация икры, подращивание личинок, выращивание молоди пресноводных и морских рыб, выполненных на базе унифицированных блоков (каркасная конструкция с сэндвич-панелями).
Преимущества рыбоводных заводов (предприятий) модульного типа: тиражируемость на специализированном предприятии (рис. 10); занимаемые площади сокращены за счет эффективного использования объема модуля; климатическая система и теплоизолированные панели обеспечивают поддержание внутри модуля выбираемых пользователем температурных режимов (в пределах от 0 до 400С с точностью 0,2-0,30С) вне зависимости от внешних метеоусловий (рис. 11); каркасная конструкция соответствует требованиям по обеспечению ветро-, снего- и сейсмоустойчивости; компоновка и габариты модулей позволяют обеспечить доставку их на место строительства рыбозавода любым видом транспорта (рис. 12, 13). Поставка модулей, укомплектованных оборудованием, позволяет произвести наладку и запуск в эксплуатацию в кратчайшие сроки (3-5 дней). Возможность перемещения модулей способствует снижению стоимости строительно-монтажных работ, значительному уменьшению объема работ нулевого цикла, проведению монтажа в любой сезон года. Возможность применения замкнутого цикла, терморегулирования, бактерицидной обработки воды позволяет управлять темпом развития и роста рыб, производить лечебно-профилактические мероприятия.
Модульное оборудование позволяет осуществить весь комплекс работ, причем модули могут иметь как смешанную функцию, так и разделяться по назначению: для инкубации икры, для выращивания молоди, для культивирования кормов, водоподготовки, размещения силового оборудования и др. Рыбоводное предприятие может быть использовано в комплексе с прудами, вырастными каналами, садковой базой (рис. 14, 15).

Рис. 10. Строительство блок-модуля

Рис. 11. Система термостатирования (тепло-холод) рабочего помещения

Рис. 12. Доставки автотранспортом

Рис. 13. Установка блок-модуля на железнодорожную платформу

Рис. 14. Бассейны для подращивания молоди

Рис. 15. Пруды на высокопрорницаемых грунтах
(геомембранная гидроизоляция)
[1] Вероятными причинами закрытия рыбозаводов здесь, являются политико-экономические (прим ред.).
 Морские рыбки
Морские рыбки
 Пресноводные рыбки
Пресноводные рыбки
 Террариумные животные
Террариумные животные
 Аквариумы и тумбы
Аквариумы и тумбы Каталог товаров
Каталог товаров Терарриумы
Терарриумы