М.Ю. Колобов (Московский государственный университет им. М.В. Ломоносова, ООО «Аква Лого», Москва)
Исследования, направленные на изучение гальванотаксиса животных, проводились в 2001 году на илисто-песчаной литорали Белого моря (губа Грязная, Кандалакшский залив) и в лабораторных условиях. В качестве модельного организма использовался литоральный брюхоногий моллюск Hydrobia ulvae.
1.1. Электрохимический профиль литорали.
Большие различия гидрохимических показателей (pH, соленость грунтовой воды и концентрации веществ, изменяющих направление и скорость окислительно-восстановительных процессов в морских осадках (Розанов, 1988)), а также физических условий (температура, Eh, длительность инсоляции и выветривания грунта), на разных уровнях литорали наводят на мысль о возможности возникновения и протекания между ними электрических токов различной силы и направленности, отличных по своей природе от морских теллурических токов.
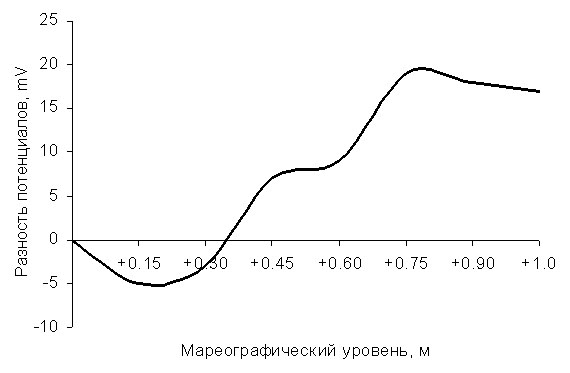
Рисунок. Разность электрохимических потенциалов Δμ между разными горизонтами литорали и морской водой («0»). Указаны значения основных физико-химических показателей в нижней и средней литорали.
Проведенные измерения позволили зафиксировать заметную разность электрохимических потенциалов (Δμ) между различными горизонтами литорали и морской водой во время отлива. Как видно на рисунке, наибольших значений Δμ достигает между морской водой и средней литоралью. Причем средняя литораль положительно заряжена по сравнению с морской водой («0»), и максимальное значение составляет 20 mV, а нижняя – отрицательно (-5 mV).
Эта закономерность, вероятно, может использоваться организмами, имеющими электромагнитную чувствительность, и их личинками как фактор ориентации во время оседания в зоне литорали и дальнейших миграций.
1.2. Поведение гастроподы Hydrobia ulvae в поле постоянного тока.
Наличие на литорали электрических токов, протекающих в поверхностных слоях грунта в определенном направлении, позволило поставить вопрос о возможности использования этого фактора бентосными и пелагическими организмами, а, возможно, и их личинками в качестве способа ориентации на пространстве литорали.
По поводу электротаксических, и, в частности, гальванотаксических реакций животных существует обширная литература. Наиболее многочисленны исследования электротропизма рыб (McMillan, 1928; Миронов, 1948; Klima, 1972; Madden, Houston, 1976; Curry, Kynard, 1978; Gatz, Loar et al., 1986; Barrett, Grossman, 1988; Barham, Schoonbee et al., 1989a; Barham, Schoonbee et al., 1989b; Bohlin et al., 1989) как пресноводных, так и морских, у которых существует ярко выраженная «анодная реакция» – движение в сторону положительно заряженного электрода (анода). Эта реакция возникает при создании вокруг рыбы электрического поля (от десятков милливольт до нескольких вольт на метр, в зависимости от вида и размера рыб) и уже несколько десятков лет используется в промышленном рыболовстве (Reynolds, Kolz. et al., 1988; Halacka, Jurada, 1994; Holliman, Reynolds, 2002). Аналогичное поведение в электрическом поле отмечено и для бентосных беспозвоночных животных (Shentyakova et al., 1970) и, в частности, ракообразного Homarus americanus (Koeller, Crowell, 1998), а также различных инфузорий (MachemerRohnisch et al., 1996; Machemer, 1998; Van Hoek et al., 1999).
Для определения влияния электромагнитного поля на поведение беломорского литорального моллюска Hydrobia ulvae было осуществлено 6 однотипных лабораторных экспериментов, каждый из которых длился 50 часов.
Общим результатом проведенных экспериментов можно признать наличие у H. ulvae переменной реакции положительно-отрицательного гальванотаксиса (ориентировка и движение в направлении одного из электродов). Проявлением такого поведения является повышение вероятности передвижения моллюсков в сторону анода или катода и дальнейшего нахождения вблизи них. Направление этих перемещений зависит от напряжения электрического тока, прилагаемого на электроды, и физиологического состояния организмов (время голодания).
Относительно небольшое напряжение (10-20 mV) между анодом и катодом вызывает «мягкий» положительный гальванотаксис – движение гидробий в сторону положительно заряженного электрода (анода), в течение которого организмы достаточно свободно перемещаются во всех направлениях, но движения в сторону анода преобладают. В результате, через 8-30 часов от начала эксперимента до 80-85% всех моллюсков может находиться вблизи анода. При достижении его некоторые особи прекращают двигаться и остаются на месте длительное время, вплоть до конца эксперимента. Другие особи в поисках пищи могут через некоторое время начать двигаться в обратном направлении, однако как и прежде, преимущественная «анодная» локализация всей совокупности моллюсков сохраняется.
Увеличение напряжения на электродах до 50 mV приводит к повышенной активности моллюсков и возникновению отрицательного гальванотаксиса – движению гидробий в сторону катода и последующей их преимущественной локализацией вблизи этого электрода. Такое распределение достигается в течение от 1 до 20 часов. Аналогичную «катодную» реакцию, хотя и менее выраженную, вызывает и длительное (более 10 суток) голодание гидробий.
Дальнейшее увеличение напряжения на электродах (до 1,5 V) приводит к интенсивному электролизу морской воды, но не вызывает электронаркоза моллюсков. Их двигательная активность сохраняется, а перемещения остаются плавными. Еще в пионерных работах по изучению реакции электротаксиса животных показано, что возникновение стадии электронаркоза зависит не только от напряженности электрического поля среды, но и от размеров тела организмов. Макмиллан (MсMillan, 1928) установил, что напряженность электрического поля, вызывающая паралич (электронаркоз) у рыб одного вида, но разных размеров обратно пропорциональна размерам рыб. Таким образом, при малых размерах тела моллюсков H. ulvae электронаркоз, вероятно, может наступить только при гораздо большей напряженности электромагнитного поля, чем создавалось в данном эксперименте, что в природных условиях практически невозможно.
Учитывая факт наличия в морском грунте разности электрохимических потенциалов и сопоставив с ним результаты данного эксперимента, можно предположить, что чувствительность к электромагнитному полю и детерминированность поведения в этих условиях могут использоваться особями H. ulvae и, возможно, другими бентосными организмами как способ ориентации.
Отмечено, что величина электрохимического потенциала верхних горизонтов литорали прямо зависит от температуры окружающей среды, интенсивности выветривания и инсоляции грунта – факторов, длительное и чрезмерное воздействие которых губительно как для самих бентосных организмов, так и для их кормовой базы (микроводоросли и мейобентос). Поэтому становится оправдано изменение положительного гальванотаксиса на отрицательный при повышенной силе электрического тока (около 50 mV), что в природных условиях может соответствовать преимущественному движению моллюсков в сторону более увлажненных и богатых органикой нижних районов литорали при неблагоприятных условиях среды в верхних районах литорали. В этом случае такое поведение моллюсков является активным механизмом защиты репродуктивного ядра популяции – крупных половозрелых особей, локализующихся преимущественно в верхних районах литорали.
Литература:
- Миронов А.Т., 1936. Об изучении электрических токов моря // Журн. геофизики. № 6. Вып. 5.
- Миронов А.Т., 1948. Электрический ток в море // Труды МГИ АН СССР. Вып. 1. С. 56.
- Розанов А.Г., 1988. Окислительно-восстановительные процессы в морских осадках и методы их исследования // В сб. «Химический анализ морских осадков». М.: Наука. С. 5-44.
- Barham W.T., Schoonbee H.J. et al., 1989a. Some observations on the narcotizing ability of electric currents on the common carp Cyprinus carpio // Onderstepoort Journal of Veterinary Research. V. 56. P. 215-218.
- Barham W.t., Schoonbee H.J. et al., 1989b. Use of electronarcosis as anesthetic in the cichlid Oreochromis mossambicus (Peters), III. The Effects of changing Physical and electrical parameters on the narcotizing ability of halfwave rectified currents // Onderstepoort Journal of Veterinary Research. V. 56. P. 67-71.
- Barrett J.C., Grossman G.D., 1988. Effects of direct current electrofishing on the mottled sculpin // North American Journal of Fisheries Managment. V.8. P. 112-116.
- Bohlin T. et al., 1989. Electrofishing - theory and practice with special emphasis on salmonids // Hydrobiologica. V. 173. P. 9-43.
- Curry K.D., Kynard B., 1978. Effect of extended galvanonarcosis on behavior of rainbow trout, Salmo gairdneri, and chanell catfish, Ictalurus punctatus. // Journal of the Fisheries Research Board of Canada. V.35. P.1297-1302.
- Gatz J.A., Loar J.M. et al., 1986. Effects of repeated electroshocking on instantaneous growth of trout. // North American Journal of Fisheries Managment. V.6. P. 176-182.
- Halacka K., Jurajda P., 1994. Characteristics of electrofishing equipment.// Zivocisna vyroba. V 39. N. 8. P 733-741.
- Holliman F.M., Reynolds J.B., 2002. Electroshock-induced injury in juvenile white sturgeon. // North Am. J. Fish Manage. V. 22. N. 2. P. 494-499.
- Klima E.F., 1972. Voltage and pulse rates for inducing electrotaxis in twelve coastal pelagic and bottom fishes.// Journal of the Fisheries Research Board of Canada. V. 26. N.11. P. 1605-1614.
- Koeller P., Crowell G.,1998. Electrotaxis in American lobsters, Homarus americanus, and its potential use in sampling early benthic-phase animals.// Fish. Bull. V. 96. N. 3 .P 628-632.
- Machemer H., 1998. Electric potentiation of gravikinesis in Paramecium is possibly mediated by filaments.// Life sciences: Microgravity research. V 21. N. 8-9. P. 1301-1309.
- MachemerRohnisch S., Machemer H., Braucker R., 1996. Electric-field effects on gravikinesis in Paramecium. // J. Comp. Physiol. A-Sens. Neural Behav. Physiol. V 179. N. 2. P 213-226.
- Madden J.A., Houston A.H., 1976. Use of electroanestesia with freshwater teleosts: some physiological consequences in the rainbow trout, Salmo gairdneri (Richardson).// Journal of Fish Biology. V.9. P.457-462.
- McMillan F.O., 1928. Electric fish screen. // Bull. US Bur. of Fish. V. 44. P. 97-128.
- Reynolds J.L., Kolz L. et al., 1988. Electrofishing injury to large rainbow trout: comments.// North American Journal of Fisheries Managment. V.8. P.516-518.
- Shentyakova L. F. et al., 1970. Effect of an alternating electric current on fishes and aquatic invertebrates // J. of Ichthyology. V. 10. N.3. P. 363-372.
- Van Hoek A.H.A.M., Sprakel V.S.I., Van Alen T.A., Theuvenet A.P.R., Vogels G.D., Hackstein J.H.P. 1999. Voltage-dependent reversal of anodic galvanotaxis in Nyctotherus ovalis. // J. Eukaryot. Microbiol. V.46. N. 4. P. 427-433.
 Морские рыбки
Морские рыбки
 Пресноводные рыбки
Пресноводные рыбки
 Террариумные животные
Террариумные животные
 Аквариумы и тумбы
Аквариумы и тумбы Каталог товаров
Каталог товаров Терарриумы
Терарриумы